




摩登7平台合作客户/
拜耳公司 |
同济大学 |
联合大学 |
美国保洁 |
美国强生 |
瑞士罗氏 |






相关新闻Info
推荐新闻Info
-
> 3种助剂对螺虫乙酯和联苯菊酯药液表面张力、金钗石斛菲盾蚧防治效果的影响(三)
> 3种助剂对螺虫乙酯和联苯菊酯药液表面张力、金钗石斛菲盾蚧防治效果的影响(二)
> 3种助剂对螺虫乙酯和联苯菊酯药液表面张力、金钗石斛菲盾蚧防治效果的影响(一)
> 利用LB膜分析仪技术制备纳米环组装阵列,得到一种具有结构色的材料
> 覆盖有疏水网的开放管道液体输运稳定性研究(下)
> 覆盖有疏水网的开放管道液体输运稳定性研究(上)
> 水、常温液态金属等9种流体对液滴碰撞壁面影响的数值研究(三)
> 水、常温液态金属等9种流体对液滴碰撞壁面影响的数值研究(二)
> 水、常温液态金属等9种流体对液滴碰撞壁面影响的数值研究(一)
> 弹簧秤测量水的表面张力系数实验装置改进措施及效果
两种抗菌肽与生物膜之间的相互作用的比较——结果和讨论、结论
来源:上海谓载 浏览 2119 次 发布时间:2022-01-12
结果
抗菌肽在脂质单层中的渗透
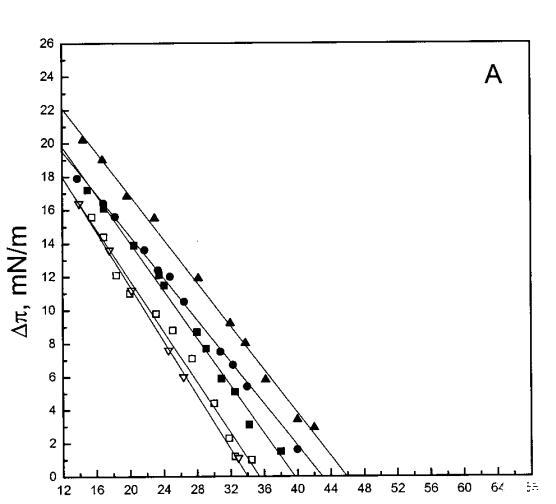
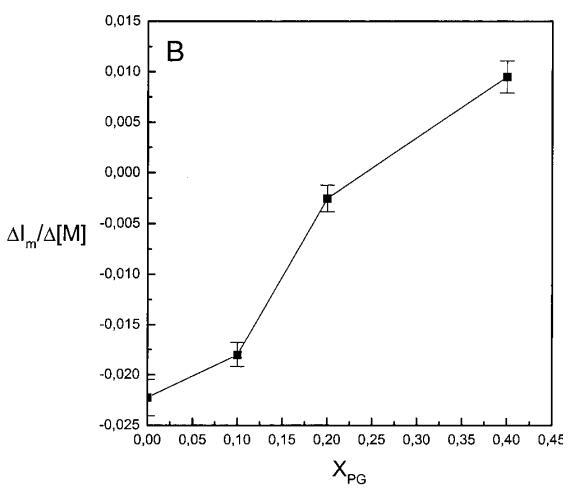
通过测量表面压力的增量,观察到抗菌肽在不同初始表面压力(π0)下嵌入到空气/水界面上的脂质单层中(∆π)将肽添加到亚相后(Brockman,1999;Verger和Pattus,1976)。Magainin 2在π0>12 mN/m时容易插入SOPC单层(图1A),而在π0以上≈35 mN/m时,其在膜中的渗透被消除,因此该值表示该肽与SOPC膜相互作用的临界包装压力πc。根据其净正电荷4(Matsuzaki,1998),当膜中酸性磷脂POPG的含量增加时,magainin 2向脂质单层的渗透逐渐增强(图1 A)。因此,消除magainin 2插入单层的πc值增加至~39(XPG=0.1)、43(XPG=0.2)和46 mN/m(XPG=0.4)(图1c)。相反,在SOPC膜中加入胆固醇(Xchol=0.1)对magainin 2的渗透没有显着影响(图1A)。
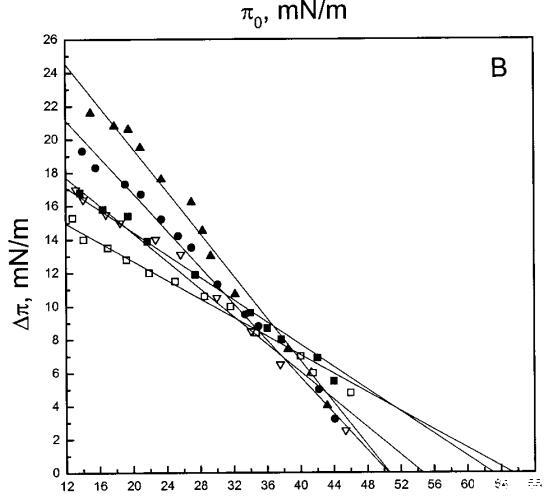
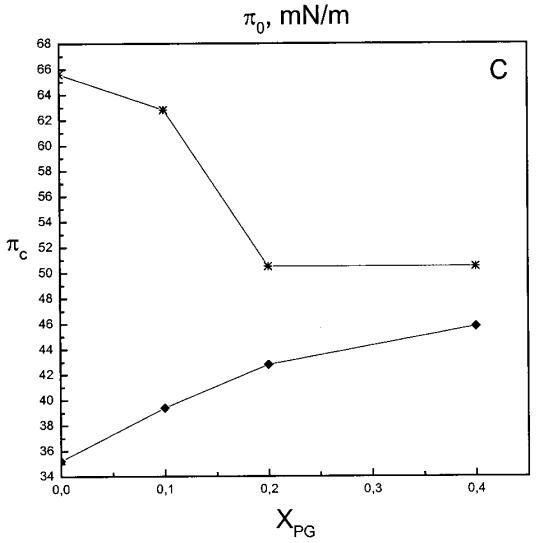

吲哚西丁的π也显着增加,显示其插入SOPC单层(图1b)。然而,吲哚青素的膜活性明显高于magainin 2,具有πc≈65mN/m,用于SOPC胶片。与magainin 2类似,单层中POPG含量的增加增强了吲哚青霉素的渗透,增加了∆π低于~38 mN/m,与肽和脂质单层之间的静电相互作用一致。然而,在后一个表面压力值之上,吲哚西丁引起的π增量被POPG减弱。因此,与magainin 2相反,在存在POPG的情况下,πc减小(图1c),在XPG=0.4时从~65 mN/m(对于SOPC单层)变为~50 mN/m。在SOPC单层中,吲哚西丁增加的π也被胆固醇(X=0.1)增加,而πc降低至~54 mN/m。图1所示的magainin 2和吲哚西丁的模式非常不同,后一种肽的明显交叉点为~38 mN/m。
抗菌肽对双层脂质动力学的影响
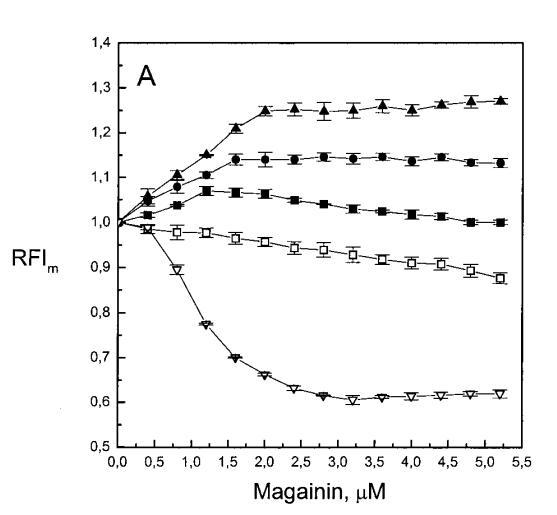
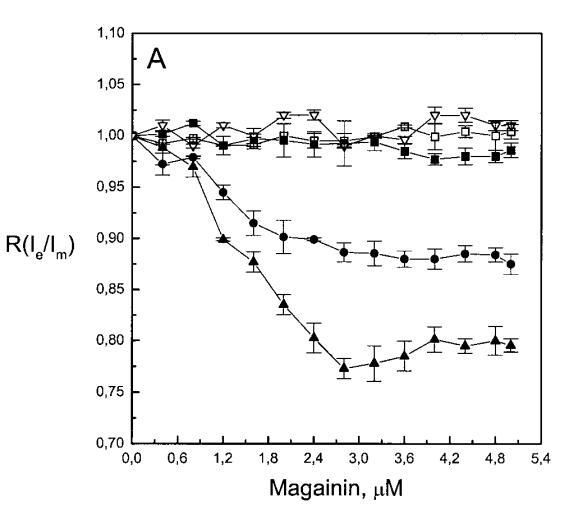
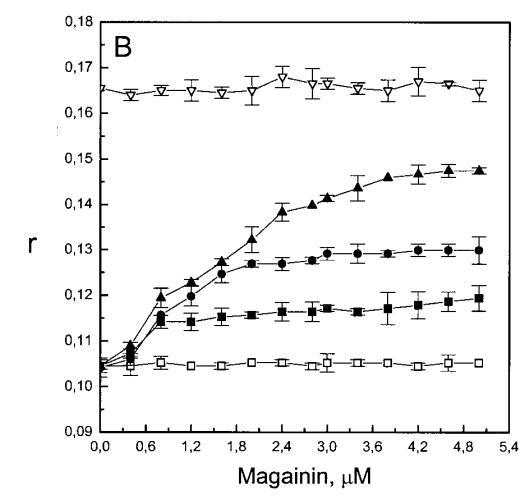
继上述单层研究之后,有兴趣探索肽对脂质动力学的影响。这是通过使用荧光光谱和不同脂质成分的LUV完成的,记录芘标记的荧光磷脂类似物PPDPC或PPDPG的发射光谱(均为X=0.01)。对于SOPC LUV,添加magainin 2导致芘荧光显着猝灭,芘单体和准分子发射强度平行降低,如Im所示(图2)。作为XPG的一项功能,还研究了马盖宁2与脂质体结合的后果。当XPG从0.1增加到0.4时,观察到由于magainin 2在398 nm处逐渐增强的Im(RFIm),而Im的初始增量在明确定义的PG/magainin 2化学计量比~2、3和5时趋于平稳(图2 A)。在这个初始阶段之后,直到给定的化学计量比,还有第二个过程,其斜率也随着XPG的增加而增加(图2b)。在XPG=0和0.1时,Magainin 2仅引起Ie/Im的轻微变化,而在XPG=0.2和0.4时,分别减少10%和25%是明显的(图3 A)。为了区分脂质动力学中两种性质不同的变化,即侧向迁移率和微结构域形成,作为上述Ie/Im变化的根本原因,我们还通过测量DPH的荧光各向异性r来评估酰基链顺序的变化(图3 B)。在缺乏酸性磷脂的情况下,Magainin 2几乎不会引起r的变化(图3B)。然而,随着XPG的增加,加入magainin 2后脂质堆积和酰基链顺序逐渐增加(图3b),XPG=0.4时,r最大增加1.4倍。值得注意的是,XPG=0.1时Ie/Im没有变化以及酰基链顺序的增加表明PPDPG正在发生微观富集。有趣的是,在存在胆固醇(Xchol=0.1)的情况下,由于马盖宁2的作用,Im和Ie都显着降低,Ie/Im的变化不显着(图2A和3A)。如前所述(van Ginkel et al.,1989),添加肽前r的增加(图3 B)揭示了通过在膜中掺入胆固醇而增强的膜顺序。然而,在胆固醇存在的情况下,magainin 2对r没有进一步的影响(Xchol=0.1,图3b)。


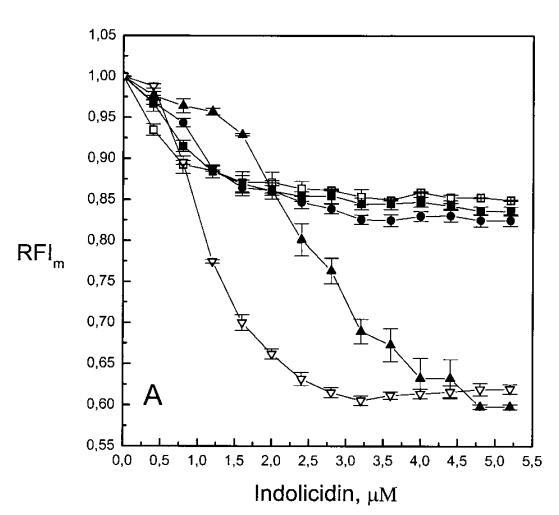
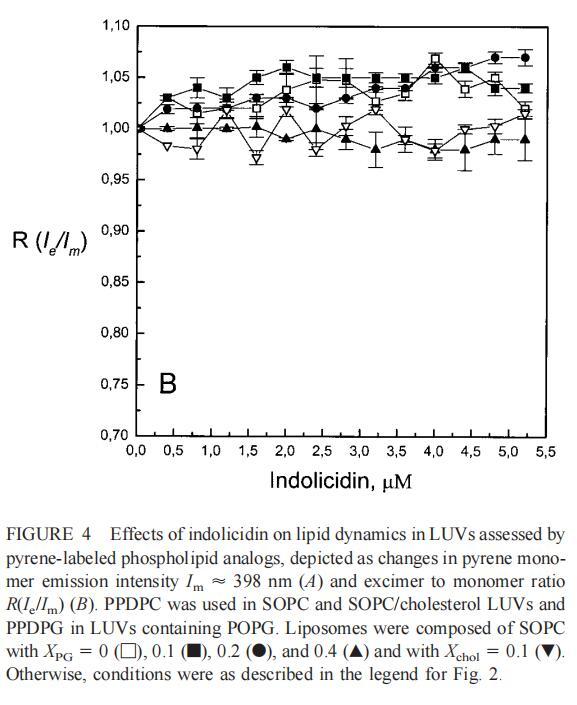
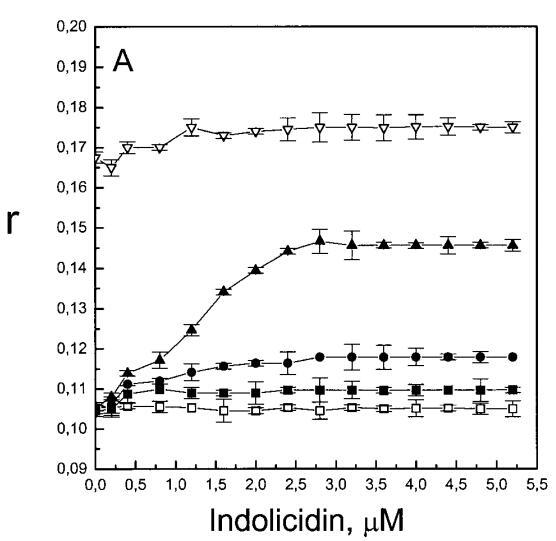
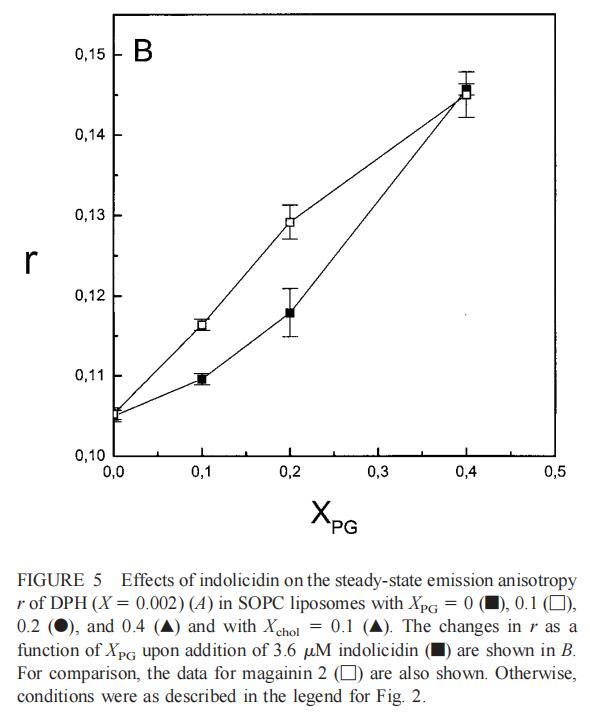
随后,我们进行了与上述相同的测量,但使用吲哚西丁。与magainin 2类似,吲哚西丁也导致SOPC LUV的Im和Ie同时减少,如图4a所示。然而,与magainin 2相反,当存在酸性磷脂POPG时,芘荧光的猝灭也很明显。当肽/脂摩尔比为1/20时,Im按XPG?0.40.20.10顺序下降。当XPG=0.4且肽/脂摩尔比>1:12.5时,Im值进一步降低。吲哚西丁引起的Im最大减量为15%(XPG=0)至40%(XPG=0.4)。尽管Im降低,但Ie/Im未显示因吲哚西丁引起的显着变化(图4b)。有趣的是,在Xchol=0.1时测量到Im的最大下降(图4a)。与magainin 2类似,当吲哚西丁添加到SOPC LUV中时,DPH各向异性r保持不变(图5a)。然而,在含有酸性磷脂的膜中,吲哚西丁导致脂质堆积增加和酰基链顺序增加,明显表现为r增加(图5a)。此外,r的增加取决于XPG,随着双层中酸性磷脂含量的增加而增加(图5b)。相应的最大增幅为1.05倍至1.39倍。在胆固醇存在的情况下,也观察到吲哚肽引起的膜酰基链顺序的增加,使r增加1.06倍(图5a)。
抗菌肽对膜拓扑结构的影响
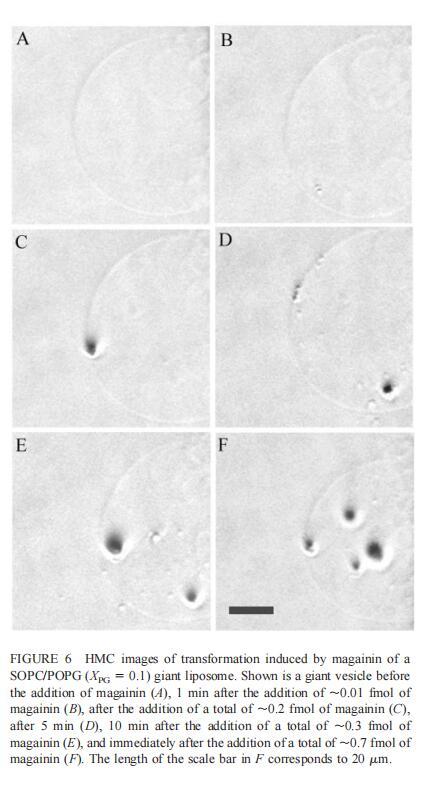
由于上述magainin 2和吲哚西啶对单层和LUV具有明显的脂质成分依赖性作用,因此有兴趣使用巨型脂质体比较这些肽对宏观膜的三维拓扑结构的影响,无论是否含有PG,都可以在交流电场中形成由SOPC组成的巨大脂质体。即使暴露于大量肽(~2 fmol,~4 pl=200次重复注射,数据未显示),SOPC膜也不受magainin 2的影响。同样,由SOPC和胆固醇(Xchol=0.1)组成的巨大脂质体也不受此量的马盖宁2的影响(数据未显示)。然而,在存在POPG(XPG=0.1)的情况下,已经~0.01 fmol(单次~20 fl注射)的magainin 2应用于巨大囊泡表面,显着影响巨大脂质体膜的拓扑结构(图6 B)。更具体地说,在~1分钟内,小颗粒(θ≈0.9米)出现在巨大的囊泡内。随后注射高达~0.1 fmol(约200 fl)的magainin 2不会引起进一步的可见变化。在总共添加约0.2 fmol的肽后,膜开始起伏,形成致密颗粒(θ≈4 m)在巨大脂质体内可见(图6 C)。在~5分钟内,几个小颗粒(θ≈0.9 m)出现在巨大脂质体膜的内表面(图6 D)。随后,大多数小颗粒进入巨大囊泡的内腔。这些内吞小泡似乎在巨大脂质体内自由移动,在随后30分钟的观察中没有进一步的变化。进一步注射magainin 2,提供总共0.3 fmol的肽,引起类似内吞作用的过程,出现直径约8 m的致密颗粒(图6 E)。约10分钟后,巨大脂质体内约0.9-m颗粒的数量继续增加。偶尔,这些颗粒也出现在巨大囊泡的外表面,并释放到外部缓冲液中(图6 E)。随后添加magainin 2(~0.7 fmol)使膜不稳定并诱导进一步的囊泡形成(图6 F)。这些内吞囊泡在巨大脂质体内彼此分离,巨大囊泡的大小明显减小(图6f)。最终,在进一步添加magainin 2(高达~1.5 fmol)后,巨大的脂质体膜崩塌。上述形态学变化具有高度的可重复性。
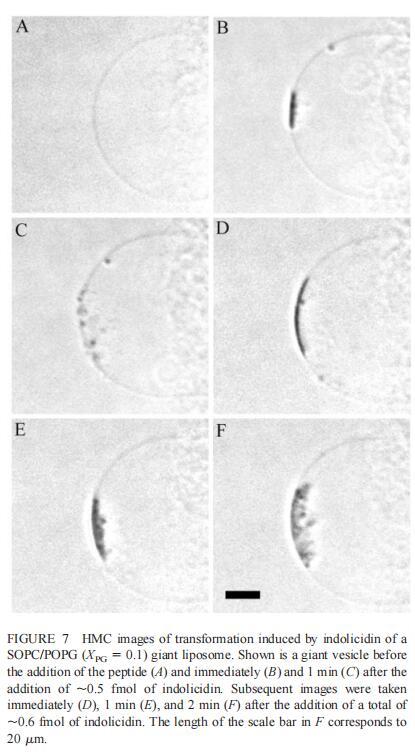
随后,我们研究了吲哚西丁对由SOPC及其与POPG(XPG=0.1)的混合物组成的巨大脂质体的影响。对于SOPC巨型脂质体,唯一明显的变化是在其表面应用~2 fmol(~200次注射)吲哚西丁后出现轻微收缩(数据未显示)。然而,当囊泡含有POPG(X=0.1)时,吲哚西丁(~0.5 fmol)诱导类似内吞作用的过程(图7),巨大脂质体内出现小颗粒(~0.9 m)。添加约0.5 fmol吲哚西丁(50次注射)后,暴露于吲哚西丁的巨大脂质体表面区域变暗,折射率发生显着变化,表明形成紧密堆积的三维结构,仅部分保留在焦平面内(图7b)。小粒子(θ)≈在上述聚集体形成后约1分钟内,巨大脂质体内出现0.9 m(图7 C)。随后再添加0.1 fmol吲哚西啶后,膜表面的聚集物仍然存在,更多的小直径颗粒出现在巨大的囊泡内(图7,DF)。这种类似内吞作用的过程在随后添加吲哚西丁的情况下重复,大量小颗粒逐渐积聚在巨大脂质体内,并且巨大脂质体的大小略有减小。
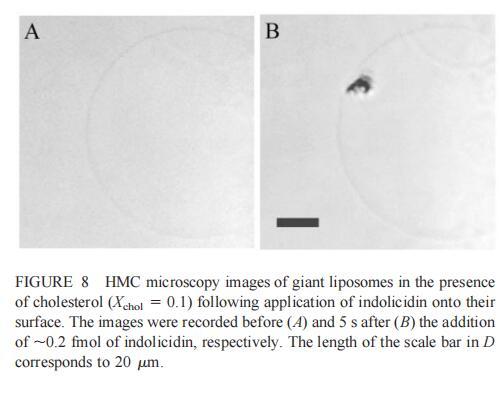
随后,我们研究了吲哚西丁对由SOPC和胆固醇组成的巨大脂质体(Xchol?0.1)的影响。吲哚西丁在胆固醇存在的情况下也会引起巨大脂质体的起泡。形成的内吞小泡在巨大小泡的表面上深度聚集(图8b)。具体地说,在添加约0.2 fmol吲哚西啶(~400 fl,20次注射,图8 B)后约5 s内,巨大脂质体表面出现一簇小颗粒。随后,聚集物转移到巨大的脂质体中,同时也有少量内吞颗粒(θ≈0.9米)处可见。进一步添加吲哚西丁会增加这些颗粒的数量,并导致巨大脂质体收缩。然而,即使在暴露于~2 fmol吲哚西丁(200次注射,数据未显示)的情况下,膜大体上仍保持完整。
讨论
马盖宁2和吲哚青霉素的膜活性
单层实验显示,magainin 2和吲哚西啶具有高度的膜活性(图1),在空气/水界面上的脂质膜下注射后,表面压力显着增加。根据膜的组成,发现了与表面压力πc对应的临界堆积密度,超过该密度,这些肽不再与单层相互作用,从而导致π的增加(Verger和Pattus,1976)。尽管这两种肽插入两性离子磷脂酰胆碱单分子膜,但它们优先与酸性磷脂相互作用,与其净阳离子电荷保持一致(Matsuzaki等人,1991、1993、1995a、1998;Ladokhin等人,1997)。然而,PG对其穿透力的影响明显不同,吲哚西丁在~38 mN/m的表面压力下有一个交叉点,吲哚西丁的斜率为∆存在PG时,π与π0的比值变得更陡(图1)。相比之下,斜坡∆?与π0相比,magainin 2的穿透力未因PG的存在而改变(图1A)。我们的数据表明,magainin 2更表面地附着于单层,其穿透深度增加PG,并随着表面压力的增加而逐渐减小,其长轴相对于层平面的角度在不同压力下保持相对不变。上述magainin 2与脂质单分子膜的相互作用模式可兼容1)其主要为=-螺旋构象,PG增加=-螺旋度(Wieprecht et al.,1997),和2)该肽在其长轴平行于层平面的情况下向脂质双层的浅层渗透(Bechinger等人,1993年;Matsuzaki等人,1994年)。在π>πc时,magainin 2仍可能与单层中的PG静电结合,尽管没有穿透膜,因此不会导致π增加。最后,我们的数据还表明,与双层不同,magainin 2不会在脂质单层中采用垂直方向(即其=-螺旋的长轴垂直于层平面)。
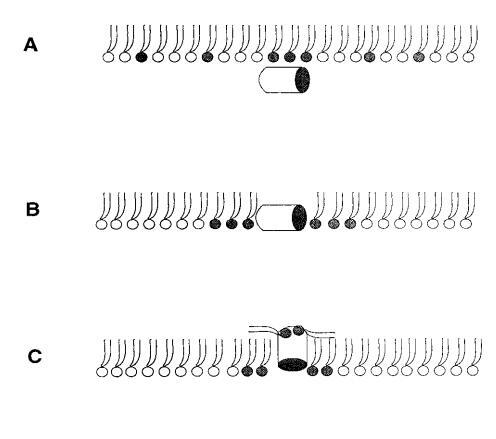
对于测试的所有单层组成,吲哚青霉素的πc值超过了马盖宁2的测量值(图1 c),表明该肽在酰基链之间有效插入和插层。吲哚西丁在~38 mN/m的交叉点值得关注。在后一个表面压力之下,表面压力的增量∆?在PG存在的情况下,吲哚西丁引起的反应增强,而在较高的初始脂质堆积密度下观察到相反的反应。有趣的是,不管脂质成分(即PG的存在)如何,交叉点几乎保持不变,因此表明它可以完全由表面压力决定,而不管静电相互作用如何。一种可能的机制是,在π0>38 mN/m时,吲哚西丁以压力依赖的方式改变其在单层中的取向,可能是从其长轴相对于层平面的平行到垂直取向(图9,B和C),从而减少面积/肽并减少∆π.此外,一些静电结合吲哚肽的PG分子也可能与单层中的磷脂方向不同。换句话说,吲哚西丁与其相关的PG将形成一种超分子复合物,其取向将取决于压力。这在图9中示意性地示出。重要的是,所提出的肽连接磷脂的重新定向将与最近提出的模型兼容,在该模型中,穿过双层的孔表面的一部分将由脂质头基构成(Ludtke et al.,1996)。
magainin 2和吲哚西啶对双层脂质动力学的影响
虽然荧光光谱显示,马盖宁2和吲哚西啶对脂质动力学的影响存在明显差异,但也存在一些明显的相似之处。因此,DPH各向异性r的增加表明,在存在酸性磷脂的情况下,这两种肽都会导致酰基链顺序的增加,这种影响的程度随着XPG的增加而增加(图3b和5b)。同样,在胆固醇存在的情况下,两种肽对芘荧光的有效猝灭也很明显(图2a和4a)。这可能与胆固醇的OH基团和马盖宁的E19残基之间的相互作用有关(Tytler等人,1995年),该肽在膜中具有跨膜取向。当Xchol=0.1时,r值的增加是由于吲哚西丁所致,而马加宁则无影响。这表明,在胆固醇存在的情况下,吲哚肽可能采用跨双层取向,其长轴垂直于双层平面。
这两种抗菌肽对芘标记的磷脂类似物的猝灭机制令人感兴趣。可能涉及π-阳离子相互作用(Dougherty,1996),后者由肽的碱性残基表示。吲哚西丁具有较高的Trp含量(39%),可促进其分配到脂质双层(Wimley和White,1996)。因此,芘和色氨酸之间的π-相互作用也是可能的。已经证明,色氨酸可以淬灭胶束溶液中各种芘衍生物的荧光发射(Encinas和Lissi,1986;Altamirano等人,1998)。另一种可能性是链反转(Holopainen et al.,1999),通过肽与脂质双层结合,导致芘部分调节到更极性的环境中,从而增加荧光团微环境的极性,并导致量子产率降低。
除上述相似性外,当使用芘标记的脂质类似物研究酸性磷脂对脂质动力学的影响时,两种肽之间也存在明显差异。Magainin 2导致两性磷脂组成的LUV的芘发射显着猝灭(图2A)。然而,在PG存在的情况下,芘的单体强度增加,XPG逐渐增加了这种效应的幅度(图2A)。圆二色性、拉曼光谱、傅里叶变换红外光谱和固态核磁共振研究表明,马盖宁在与脂质双层结合时采用螺旋构象(Matsuzaki等人,1989年、1991年;Williams等人,1990年;Jackson等人,1992年;Bechinger等人,1993年;Wieprecht等人,1997年;Hirsh等人,1996年)。magainin 2对两种膜的这些不同影响可能是由于1)通过与肽的带正电基团(例如赖氨酸的?氨基)直接接触使芘荧光猝灭,或2)插入双层的=-螺旋肽改变荧光团的微环境,与层平面平行,如上所述。然而,在PG存在的情况下,肽的正电荷应在与酸性磷脂形成静电复合物时被中和,从而减少由于π-阳离子相互作用导致的芘荧光猝灭。
有趣的是,在存在酸性磷脂的情况下,肌动蛋白/脂质的摩尔比(在该摩尔比下,肌动蛋白的初始增量趋于稳定)分别为1:21、1:16和1:12.5,对应于XPG=0.1、0.2和0.4(图2A),分别产生1:2、1:3和1:5的肌动蛋白/磷脂化学计量比。同时,DPH的初始r增量在magainin/脂质摩尔比为~1:30、1:12和1:5时趋于平稳(XPG分别为0.1、0.2和0.4),对应于大约1:3、1:2.5和1:2的magainin/PG化学计量比(图3)。据报道,在这些肽/脂质摩尔比下,magainin可插入含酸性磷脂的膜中(Ludtke等人,1994年)。此外,已经证明,双分子层中magainin的=-螺旋的方向取决于肽与脂质的摩尔比,在上述化学计量比下,螺旋应垂直于膜表面插入,在含PG的膜中形成孔(Ludtke等人,1996)。这些孔不仅由两亲螺旋的极性面组成,而且还由脂质的极性头基组成,内外单层通过这些孔衬脂质是连续的。膜中的这种结构可能会降低探针芘部分微环境的极性,从而提高其量子产率。有趣的是,当XPG>0.1时,magainin 2逐渐降低Ie/Im值(图3 A),这与增强的酰基链顺序减少脂质侧向扩散的速率一致。然而,由于magainin 2已经在XPG=0.1时增加了r,Ie/Im没有变化,我们可以得出结论,PPDPG探针向微区富集以及其横向扩散的减少正在发生。
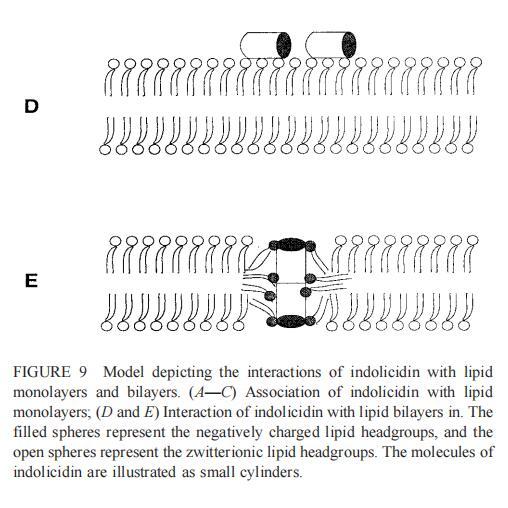
与magainin 2类似,吲哚青霉素与阴离子和两性离子囊泡的膜结合非常不同(图1b、4和5),与先前的平衡透析研究一致(Ladokhin et al.,1997)。虽然芘荧光的猝灭显示吲哚肽被分配到SOPC双层中,但对膜酰基链顺序几乎没有影响(图5)。相反,吲哚西啶与阴离子膜的相互作用导致脂酰链顺序增加,明显表现为DPH荧光各向异性增加,并对XPG呈S形依赖(图5)。与magainin 2相反,吲哚西丁导致芘单体和准分子发射强度显着平行降低,与脂质成分无关,Ie/Im变化较小(图4)。相应地,吲哚西丁诱导酸性PG富集到微区。有趣的是,在吲哚西丁/脂质摩尔比为1:20以下,芘Im的减少顺序为XPG=0.40.20.10(图4a),而在吲哚西丁/脂质摩尔比为1:10时,Im的减少对XPG的依赖性相反。这种变化的可能机制如下。在低肽/脂摩尔比时,吲哚西丁似乎被调节到双层界面(Ladokhin等人,1997),芘标记链的链反转(Holopainen等人,1999)使该部分更接近双层表面,从而允许其与吲哚西丁的基本残基接触。后者通过π-阳离子相互作用导致芘部分的碰撞猝灭。然而,随着XPG的增加,我们可以预期PG头基与吲哚西丁的碱性残基发生静电结合,从而阻止芘与吲哚西丁的阳离子残基之间的直接接触。随着肽浓度的增加,吲哚肽的膜结合以协同方式增强,猝灭效率的逆转可能涉及双分子层中脂肽复合物的聚集。因此,在酸性含磷脂的膜中,吲哚对芘的猝灭作用较小。当吲哚肽的浓度增加到肽/脂比1:10以上时,肽将以高度协同的方式并入膜的酰基链区域(Ha等人,2000)(图9 E),而在中性膜中,肽位于界面(图9 D)。插入吲哚西丁的量取决于双层中的XPG,导致吲哚西丁的Trp残基的猝灭占主导地位,与XPG=0.4时Im的显着减少一致(图4a)。
马盖宁2和吲哚西啶对巨脂质体的影响
细胞质膜通透性被认为是马加宁和吲哚肽等肽的抗菌作用的原因(Matsuzaki,1998,1999;Sitaram和Nagaraj,1999),并提出了几种机制模型(综述见Bechinger,1999)。孔或通道的形成被认为会导致跨膜电位的耗散和脂质不对称(Falla等人,1996年;Matsuzaki,1998年,1999年;Sitaram和Nagaraj,1999年)。Magainin 2和吲哚肽应按照定义的结构插入脂质双层,与所有疏水氨基酸形成构象,形成一个表面,然后与膜的疏水核心接触。因此,这些肽的膜插入迫使磷脂酰基链分开。这些肽的抗菌作用还可能包括局部相变,如脂肽胶束从双层分离,或由于插入肽引起膜曲率的局部变化,这可能导致双层结构的局部坍塌。在分子水平上,这些过程可能是本文报道的magainin 2和吲哚美辛诱导的巨大囊泡形成的基础。magainin的裂解活性对靶膜的脂质成分高度敏感,该肽优先结合并渗透阴离子脂质膜(Matsuzaki等人,1991、1993、1995a、1998)。因此,对微生物致死浓度的马盖宁不会裂解哺乳动物细胞,因为哺乳动物细胞的质膜外层不含带负电的磷脂(Ma tsuzaki,1999)。大肠杆菌暴露于magainin会导致细菌表面出现气泡(Matsuzaki等人,1997年)。在本研究中,观察到由马盖宁2和吲哚青霉素诱导的巨大脂质体的囊泡形成,这代表了它们与脂质双层结合的更宏观的变化。重要的是,酸性PG的存在是两种肽产生囊泡所必需的。巨大脂质体的囊泡形成需要反复应用magainin 2,因此暗示了一种浓度依赖性作用机制。已经证明,在存在magainin的情况下形成的孔隙是短暂的,具有有限的寿命(Matsuzaki等人,1995b)。当附近的孔隙崩解时,可能形成肽和脂质的超分子复合物,从而导致双层的短暂破坏(图6)。同时,一些肽分子应该转移到膜的内部小叶中(Matsuzaki等人,1995a,1996),从而使膜重组和愈合。由于magainin 2而形成的内吞小泡比胞吐更明显(图6),这可能与肽在双层外表面的不对称应用有关。因此,巨大脂质体的囊泡化可以缓解由于不对称肽结合和随后的膜膨胀而导致的双层不利应变。囊泡化可以降低剩余巨大脂质体膜中的肽浓度,从而减少孔隙形成。随后,需要继续向膜中添加magainin 2以引起巨大脂质体的进一步囊泡化(图6)。
吲哚西丁只含有13个氨基酸,因此太短,无法跨越双层。然而,该肽可以作为聚集物或带有相关酸性磷脂的头尾二聚体横跨双层(图9 E),从而导致膜不稳定性。已经证明,吲哚西丁能够通过穿过外膜杀死革兰氏阴性细菌,并通过形成通道破坏细胞质膜(Falla et al.,1996)。与magainin 2的作用相比,吲哚青霉素诱导的内吞小泡较小(图7)可能反映了这些肽与脂质相互作用的不同模式。一些吲哚肽分子可能会转移到膜的内部小叶中。除了具有广谱的抗真菌和抗菌活性外,吲哚西丁对哺乳动物细胞也有毒性(Schluesener等人,1993年;Ahmad等人,1995年)。有趣的是,吲哚西丁对SOPC和SOPC胆固醇巨型脂质体的作用不同。在这一阶段,根本原因尚不确定,可能与胆固醇对双层结构以及吲哚肽在膜中的取向的影响有关。因此,需要进一步的研究来阐明胆固醇对吲哚肽膜结合的影响。
我们的结果表明,在带负电荷的磷脂存在下,magainin 2对所用生物膜模型的作用显着增强。除了膜负电荷外,肽/脂质的化学计量也被证明是双层中马盖宁组织的重要决定因素(Ludtke等人,1994)。在低肽/脂质比率下,=-螺旋肽基本上平行于膜表面,并浅穿透膜的疏水核心(Bechinger等人,1993年;Matsuzaki等人,1994年)。该位置将脂质头基推到一边,迫使疏水区域形成间隙,而膜变薄以填充间隙(Ludtke et al.,1995)。先前已强调了由magainin形成的孔隙的瞬时性质(Matsuzaki等人,1995b)。膜的局部和瞬时破坏可能需要肽与靶细胞膜的不对称结合。因此,在magainin与靶细胞质膜双层的外小叶结合后,该外单分子层的侧压力比内小叶增加。此外,平行于层平面的=-螺旋的表面位置在外层单层中施加曲率应变,导致正的自发曲率。当两个小叶之间的侧压差大小和与正自发曲率相关的能量超过膜变形所需的能量时,双分子层不稳定,肽相对于层平面的垂直方向发生重新定向,孔隙打开。这些孔随后可能在靶体内聚集成超分子结构,如在巨型脂质体中观察到的(图6)。因此,孔隙和不稳定状态将是暂时的,允许双层愈合。为此,马格蛋白2在其序列的中间含有一组正电荷。同样,当该肽在双层中形成瞬时孔并垂直于膜平面定向时,这些阳离子残基也可能静电结合到PG,类似于此处提出的吲哚肽组织(图9)。原核细胞和真核细胞膜脂质成分的主要区别在于后者富含胆固醇(Op den Kamp,1979)。上述机制也将与胆固醇的存在相兼容,以防止magainin 2的膜扰动作用。更具体地说,胆固醇在人类质膜的两个小叶之间转移(触发器)非常快(Lange et al.,1981)。因此,这种胆固醇的快速易位可以消除在magainin 2结合到后一单层后靶膜内外小叶侧压力的差异。因此,与细菌靶点相比,这种肽在真核细胞中产生毒性作用需要大量的magainin 2(Zasloff,1987)。据估计,真核细胞质膜外小叶的平衡侧压力约为32 mN/m(Demel等人,1975年)。在超过34 mN/m的压力下,观察到吲哚西丁高效渗透到单层中,这可能部分解释了该肽在真核细胞中的溶解作用。此外,哺乳动物质膜的外小叶仅由两性离子磷脂组成,而细菌膜含有大量带负电的PG(Op den Kamp,1979)。因此,由于缺乏负电荷和高压,预计magainin 2与真核细胞质膜的相互作用较弱,而更有效的插入吲哚肽将能够干扰真核细胞双层的组织。因此,脂质特异性可能导致这两种肽的不同毒性。
结论
由于抗菌肽的详细作用机制尚待阐明,比较两种结构上不相关的肽吲哚肽和马盖宁2对模型生物膜的影响很有意义。尽管它们的构象不同,且前者的长度约为后者的一半,但它们也具有诸如净正电荷和两亲性等特征,导致它们与磷脂膜结合。酸性磷脂进一步增强了这种相互作用,这可能对它们的抗菌活性很重要。酸性磷脂是这些肽囊泡形成巨大脂质体所必需的,它们都会导致酸性磷脂分离,将它们络合成超分子簇。同样,这两种肽都增加了酰基链顺序。所有这些作用都与抗菌作用所必需的协同膜结合相一致,可能涉及肽的瞬时跨膜定向。还可能是,它们的抗菌作用需要目标膜最初不对称地暴露于这些肽中,并与系统放松回到热力学平衡有关。